
| Abbikine产品:Anti-plant actin mouse monoclonal antibody (A01050, Abbkine, Wuhan, China) 英文名:Fungal cellulase is an elicitor but its enzymatic activity is not required for its elicitor activity 中文名:真菌纤维素酶是一种激发子,但其激发子活性并不需要其酶促活性 作者单位:山东农业大学植物病理学系 研究方向:植物病理学 杂志名称:MOLECULAR PLANT PATHOLOGY (2015) 影响因子:12.19 |
研究背景
广泛的土壤传播病原体茄根枯萎病菌是一种担子菌真菌,能够感染许多植物,包括许多重要的商业作物,如玉米,水稻,小麦,马铃薯和棉花。 它主要通过形成穿透植物组织的感染钉来感染植物。 在感染过程中,这种真菌会分泌细胞壁降解酶(CWDE),例如角质酶,果胶酶,木聚糖酶和纤维素酶。
纤维素是大多数植物中主要的细胞壁成分之一。 纤维素酶催化纤维素中β-1,4-糖苷键的降解。 它们分为三种类型:内切葡聚糖酶,外切葡聚糖酶和β-葡萄糖苷酶。 内切葡聚糖酶随机水解内部糖苷键,导致聚合物长度迅速减少,糖浓度降低逐渐增加。 外切葡聚糖酶通过从还原端或非还原端去除纤维二糖来水解纤维素链,导致还原糖快速释放,但聚合物长度几乎没有变化。 内切葡聚糖酶和外切葡聚糖酶在纤维素上协同作用产生纤维二糖,然后被β-葡萄糖苷酶切割成葡萄糖。
许多植物致病真菌产生纤维素酶。但是,与其他CWDE(如角质酶,果胶酶和木聚糖酶)相比,纤维素酶在致病性中的作用鲜为人知。纤维素酶对植物致病真菌致病性的重要性尚不清楚。已证明一些纤维素酶与致病性有关。有人提出,菜豆巨噬菌利用内切纤维素酶来致病。纤维二糖水解酶在Secale谷类食品上的紫氏克拉维菌的初始感染阶段表达。植物病原真菌链格孢产生一种内切纤维素酶,这是柿子果实疾病发展的重要因素。在甘蓝型油菜和芥菜子叶和叶片的感染过程中,可检测到黄斑狼疮杆菌纤维二糖水解酶的转录。
然而,纤维素酶基因的破坏几乎总是未能表明它对致病性至关重要。玉米病原菌Cochliobolus carbonum中外切葡聚糖酶基因的破坏不会影响玉米的致病性。灰葡萄孢菌的内切葡聚糖酶基因(cel5A)的破坏导致菌株具有与番茄叶片上的野生型(WT)相同的致病性。真菌纤维素酶系统本质上非常复杂。大多数研究表明,真菌会产生多种形式的纤维素酶成分。具有相似活性的单个纤维素酶的多样性可能是大多数基因敲除实验不支持单个纤维素酶在真菌致病性中的重要作用的原因。
许多植物致病真菌产生纤维素酶。但是,与其他CWDE(如角质酶,果胶酶和木聚糖酶)相比,纤维素酶在致病性中的作用鲜为人知。纤维素酶对植物致病真菌致病性的重要性尚不清楚。已证明一些纤维素酶与致病性有关。有人提出,菜豆巨噬菌利用内切纤维素酶来致病。纤维二糖水解酶在Secale谷类食品上的紫氏克拉维菌的初始感染阶段表达。植物病原真菌链格孢产生一种内切纤维素酶,这是柿子果实疾病发展的重要因素。在甘蓝型油菜和芥菜子叶和叶片的感染过程中,可检测到黄斑狼疮杆菌纤维二糖水解酶的转录。
然而,纤维素酶基因的破坏几乎总是未能表明它对致病性至关重要。玉米病原菌Cochliobolus carbonum中外切葡聚糖酶基因的破坏不会影响玉米的致病性。灰葡萄孢菌的内切葡聚糖酶基因(cel5A)的破坏导致菌株具有与番茄叶片上的野生型(WT)相同的致病性。真菌纤维素酶系统本质上非常复杂。大多数研究表明,真菌会产生多种形式的纤维素酶成分。具有相似活性的单个纤维素酶的多样性可能是大多数基因敲除实验不支持单个纤维素酶在真菌致病性中的重要作用的原因。
目前,一些确凿的数据支持纤维素酶参与发病机理,但有关纤维素酶作为植物与病原体相互作用的诱因的信息很少。由于纤维素酶在1988年被报道为引发剂,因此只有很少的研究调查了纤维素酶的引发剂活性。在这些研究中,纤维素酶来自非致病性木霉属。被证明能诱导倍半萜类植物抗生物素蛋白的生物合成和辣椒素在植物中的积累。但是,尚不清楚纤维素酶如何在植物与病原体的相互作用中起诱导作用。特别地,纤维素酶的激发子活性是否参与其酶活性尚不清楚。在这里,我们报道了来自R. solani的内切葡聚糖酶(EG1)的克隆,定点诱变和异源表达。使用激发子反应的分析来研究激发子与植物的相互作用,以及突变分析和马铃薯X病毒(PVX)表达系统,我们证明了EG1是一个激发子,但其激发子活性独立于其酶促活性。
实验目的
植物病原性真菌产生纤维素酶。但是,关于纤维素酶作为植物与病原体相互作用的诱因的信息很少。在此,从茄红枯萎病菌中分离出一种内切纤维素酶(EG1)。它包含一个227个氨基酸的推定蛋白,带有信号肽和45族糖基水解酶结构域。 32位的天冬氨酸(Asp)残基变为丙氨酸(Ala),导致其催化活性完全丧失。内切葡聚糖酶的野生型和突变形式在酵母中表达并纯化至同质。纯化的野生型和突变体形式诱导了玉米,烟草和拟南芥叶片中的细胞死亡,以及玉米和烟草中三个防御标记基因的转录以及与玉米防御反应相关的10个基因。
此外,它们还诱导了悬浮培养的烟草细胞的活性氧(ROS)积累,中等碱化,Ca2 +积累和乙烯生物合成。同样,使用马铃薯X病毒(PVX)表达系统在烟草中产生EG1野生型和突变形式可引起细胞死亡。在体内,EG1的表达还与玉米被茄红吸虫感染期间的细胞死亡有关。这些结果提供了直接的证据,证明内切葡聚糖酶是引发剂,但是其酶活性不是其引发剂活性所必需的。
材料与方法
- 植物种植:在标准温室条件下种植玉米(Zea mays)植物(来自中国山东的农大108,是中国栽培最广泛的杂交种)和烟草植物(来自中国烟草种业有限公司的烟草Nicotiana cv。NC89)。 在25°C下,光照12小时和黑暗12小时的周期。 拟南芥植物在20–21°C的光照下生长8小时。
- 菌株、质粒和培养基
- 真菌培养
- 烟草细胞的悬浮培养
- 酶分析和蛋白质测定
- cDNA克隆
- 定向诱变
- 表达系统构建
- SDS-PAGE
实验结果
1.内切葡聚糖酶基因的克隆和序列分析。
根据我们先前在2009年从solani AG-1-IA克隆的内切葡聚糖酶基因(GU372728)的核苷酸序列,我们获得了内切葡聚糖酶基因的开放阅读框(ORF)cDNA(称为Eg1)。 内切葡聚糖酶基因的ORF编码具有227个氨基酸残基的推定蛋白质(称为EG1)。 使用SignalP(http://www.cbs.dtu.dk/service/SignalP),潜在的信号肽被预测为氨基酸1–20,表明内切葡聚糖酶是一种分泌的酶。 成熟的内切葡聚糖酶由207个氨基酸组成,计算的分子量为23.66 kDa(http://www.expasy.ch)。 BLASTP搜索的内切葡聚糖酶的假定氨基酸序列表明,它属于45族糖基水解酶(GHs)。 Zheng等人(2013)报道,茄根枯萎病菌EG1与茄根枯萎病菌内切葡聚糖酶(ADV02787)具有100%的同一性。
2.WT和突变体的EG1在毕赤酵母中的表达
使用定点诱变,将WT形式Eg1的32位天冬氨酸(Asp,D)残基编码的密码子GAC更改为编码丙氨酸(Ala,A)的GCC以产生WT,D32A的突变形式 。 EG1的WT和突变形式在酵母中表达,并纯化至十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)均匀。 用SDS-PAGE纯化的WT和EG1突变形式的分子量约为28.0 kDa(图1A)。 该值接近由内切葡聚糖酶的推导氨基酸序列估计的23.66kDa。 通过SDSPAGE,在用空质粒pPIC9K转化的酵母的培养滤液中的相应蛋白质制品(用作对照样品,CK)中未发现可见的蛋白质条带。 与野生型相比,EG1的突变形式丧失了所有的酶活性,而CK没有酶活性(图1B)。
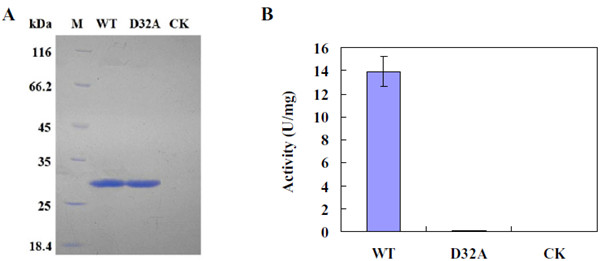
图1(A)茄红假单胞菌EG1纯化酶的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)
3.WT和EG1突变形式诱导的诱导应答
引发子在植物中诱导细胞死亡是很常见的。 将0–100 nM浓度的纯化的WT1和突变型EG1注入玉米叶片以测试其诱导细胞死亡的能力。 WT和EG1的突变形式诱导了玉米,烟草和拟南芥叶片的细胞死亡(图2A,C,D)。 浓度为40 nM的EG1接种后3天,玉米叶片出现明显症状。 注射CK不会在玉米叶片上产生任何可见的症状。 此外,使用镍亲和色谱法从用质粒pPIC9K / C61F,pPIC9K /转化的酵母的培养滤液中纯化嗜热毛毛球菌的纤维素酶(C61F)和锰超氧化物歧化酶(SOD)以及金黄色嗜热菌的纤维素酶(T61F)。 T61F和pPIC9K / SOD(图S2A–C,请参阅支持信息)。 将三个不相关的蛋白质样品C61F,T61F和SOD注入玉米叶片中,并且不会诱导细胞死亡(图2B)。
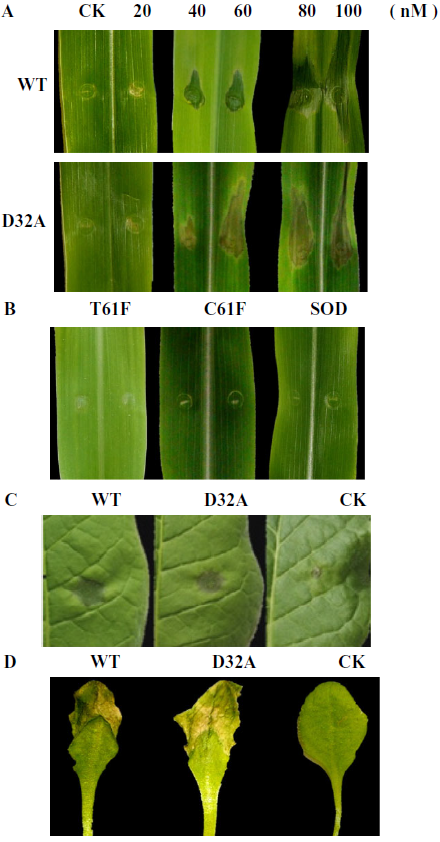
图2 茄红枯萎病菌EG1和其他真菌产生的不相关酶诱导植物叶片死亡。
4.茄红根瘤菌EG1诱导防御反应基因研究
正如已经证明的,EG1可以诱导玉米和烟草叶片的细胞死亡,我们通过逆转录-聚合酶链反应(RT-PCR)研究了EG1处理后这些植物中防御标记基因的表达模式。这些标记基因包括与发病相关的1a(PR1a)基因,过氧化物酶(POD)基因和苯丙氨酸解氨酶(PAL)基因.WT和EG1突变体形式明显诱导了玉米和烟草叶片中PAL,POD和PR1a的更高表达比做CK(图3A)。除了上述三个防御标记基因外,玉米基因组中与防御反应相关的10个选定基因的表达,例如几丁质酶,β-1,3-葡聚糖酶,发病相关蛋白,替代氧化酶,索马甜和谷胱甘肽转移酶通过实时定量RT-PCR(RT-qPCR)分析。分析表明,这10个基因在WT和EG1突变形式的玉米叶片中表现出相似的上调表达模式(图3B)。
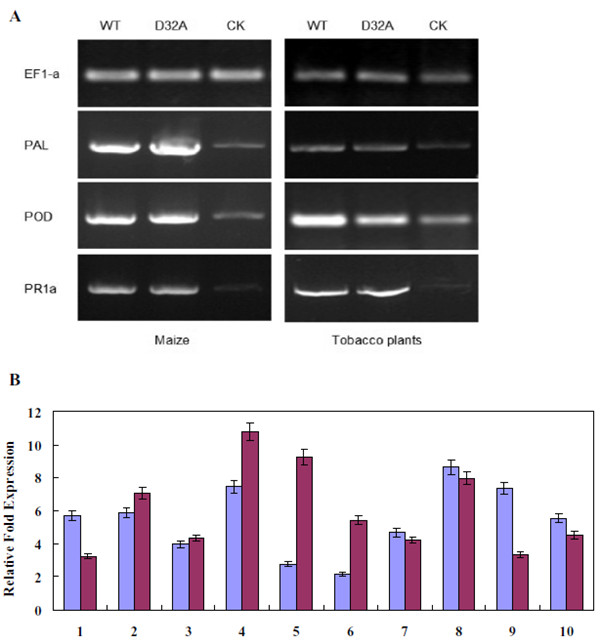
图3 茄红根瘤菌EG1诱导防御反应基因。
创新点
已经出现了一种模型,其中病原体相关分子模式(PAMP)是与宿主中的模式识别受体(PRR)识别的病原体组相关的分子。 在植物中,PAMPs作为引发剂激活固有的免疫反应,保护植物免受感染。 迄今为止,已经从多种植物致病性和非致病性微生物中分离了许多具有各种结构的蛋白质引发剂,例如鞭毛蛋白,延伸因子,转谷氨酰胺酶,木聚糖酶,纤维素结合引发剂凝集素,harpin和诱导素。 然而,迄今为止,几乎没有关于纤维素酶作为引发剂的详细信息。 我们的结果表明,真菌R. solani的纤维素酶EG1能够诱导细胞死亡,防御基因的转录,ROS的产生,中等碱化,Ca2 +积累和乙烯的生物合成,表现出代表性蛋白质激发子的特征。
还不了解植物细胞如何识别EG1。我们假设EG1的激发子活性可能基于EG1激发子与植物中可能的潜在蛋白受体之间的特异性识别。已经证明植物已经通过存在PRR而进化为识别PAMP。已鉴定出植物中的几种PRR,例如FLAGELLIN-SENSING 2(FLS2),延伸因子Tu受体(EFR)(Zipfel等,2006),LeEIX1 / 2(Ron和Avni,2004)和对芽孢杆菌多聚半乳糖醛酸酶的反应1( RBPG1)。 FLS2是最早被鉴定和研究最深入的PRR,并且是一种代表富亮氨酸的重复受体样激酶(LRR-RLK)。在拟南芥中,FLS2与相关的LRR-RLK BAK1有助于抵抗细菌病原体,BAK1可以正向调节FLS2的功能。 EFR是FLS2的紧密同源物,属于LRR-RLK的同一子小节XII。 LeEIX1 / 2是类似于番茄Ve抗性基因的LRR受体样蛋白(LRR-RLP)。 RBPG1是识别真菌内多半乳糖苷酶的拟南芥属LRR-RLP。我们认为,这些当前已知的PAMP受体将为鉴定EG1可能的潜在蛋白受体提供一些线索。
Abbkine专注于免疫学和细胞学领域,致力于创新和研发各类抗体、蛋白质、分析试剂与试剂盒,以期成为生命科学研究发展、药物研发等领域的关键推动者。在这里,我们为您呈献蛋白和免疫研究用户最喜爱的产品,从免疫学基础产品,如蛋白提取定量,到免疫学实验的内参标签抗体、一抗及二抗等;细胞研究用户最喜爱的产品,从用于检测细胞状态的染料及试剂盒,细胞器提取试剂盒,细胞亚结构染色示踪及细胞代谢检测产品,到用于细胞培养的细胞因子及蛋白类检测试剂盒,只为助力您的研究事业!
About Abbkine Scientific Co., Ltd.
Abbkine Scientific Co., Ltd.成立于2012年。我们的使命,通过为全球科学家提供人类和动物健康研究所需的基础研究工具,以期成为研究探索的关键推动者。我们将通过明确的核心战略和促进诚信、开放、合作、创新和为客户服务的企业文化,为全球生命科学研究者提供买得起的高品质产品,实现我们的使命。







