神经肽(Neuropeptides)是一类由神经元分泌的小分子蛋白质,它们在神经系统中起着调节神经传递和调节情绪、行为等功能的作用。神经肽包括多种类型,如内啡肽、去甲肾上腺素、催产素等,它们在神经科学研究中具有重要的意义。
合成与加工
神经肽由神经元合成和/或使用(Kastin, 2000, Russo, 2017)。它们最初是作为大型无活性前体蛋白质的前肽的一部分合成的(图1)。前肽含有一个N端信号肽,这是新合成的基因产物进入内质网腔所必需的。信号序列在穿过内质网膜的过程中被切断,在ER-Golgi中产生前肽,以进一步分选到受调节的分泌途径中(Burbach, 2011)。为了释放生物活性神经肽,前肽被内肽酶和外肽酶进一步切割,并在翻译后进行修饰,例如糖基化、磷酸化、硫酸化、乙酰化、添加寡糖和N端焦谷氨酸形成(Hook, 2008, Sun and Zhao, 2017, Mains and Eipper, 1999)。最常见的修饰是C端酰胺化(Eipper, 1992)。已知的生物活性肽中约有一半是α-酰胺化(图1)(Mains and Eipper, 1999)。
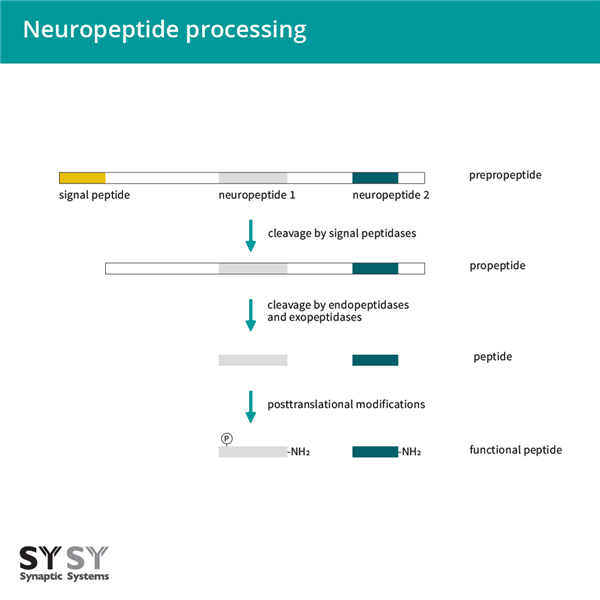
图1. 神经肽的加工过程。神经肽是作为大而无活性的前体蛋白质(前肽)的一部分合成的。几个蛋白水解切割步骤和翻译后修饰导致生物活性肽的产生。
内切蛋白水解和翻译后修饰同时发生在反式高尔基网络和多肽包装的致密核心囊泡中。 致密核心囊泡在神经元中传输,可以在突触间隙、细胞体和轴突上释放多肽(Mains and Eipper, 1999, Russo, 2017)(图2)。
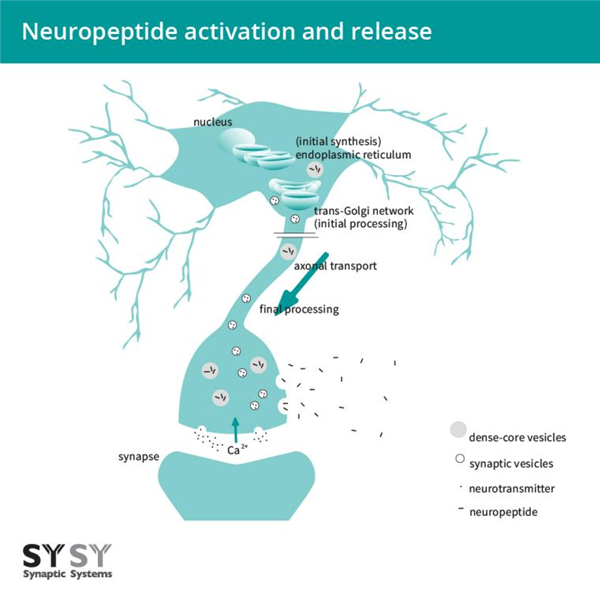
图2:神经肽的激活和释放。神经肽最初在内质网合成,在跨高尔基网络和多肽包装的致密核心囊泡中被切割和翻译后修饰。神经肽在致密核心囊泡中沿轴突运输,在Ca 2+ 流入时释放,并通过扩散分布。
神经肽的释放、分散和失活
神经肽经常与其他神经肽和神经递质一起在单个神经元中共同释放,导致各种效应(Hökfelt等,2003年,van den Pol,2012年)。在突触中,密集核囊与含有谷氨酸等经典神经递质的突触囊泡共定位(Russo,2017年)。虽然密集核囊和突触囊泡经常共同释放,但它们使用不同的机制。像神经递质一样,神经肽在去极化或其他信号的作用下,通过钙依赖的胞吐释放(Russo,2017年)。然而,与突触囊泡中的神经递质不同,密集核囊中的神经肽在胞质[Ca2+]浓度较低时释放。传统神经递质的释放被认为发生在Ca2+进入的位置附近,而神经肽通常在离Ca2+进入的位置一定距离处释放。因此,相对于Ca2+进入的位置,密集核囊的位置可能决定了分泌所需的Ca2+量(Mains和Eipper,1999年)。
与经典的神经递质不同,神经肽从释放部位扩散,因此可以在相对较大的距离(nm到mm)上发挥作用(图2)。这种扩散驱动的分布被称为体积传输或分散(van den Pol, 2012, Russo, 2017)。由于肽没有再摄取机制,它们只能慢慢地从细胞外空间移除。相比之下,经典的神经递质通过专门的转运蛋白迅速从突触间隙移除。体积传输和缺乏再摄取的结合有助于神经肽的相对持久效应(Russo, 2017)。与神经递质相比,神经肽的寿命较长,但其作用终止。细胞外蛋白酶会使其失活,在某些情况下,细胞外蛋白酶甚至可以通过裂解现有的神经肽来产生新的生物活性肽。(Russo, 2017)。
受体激活
所有神经肽都通过细胞表面受体作为信号传导器。几乎所有的神经肽都作用于G蛋白偶联受体,触发第二信使级联来调节细胞活性(Hökfelt et al., 2003, Russo, 2017)。与肽配体一样,受体不仅广泛分布在神经系统中,而且还分布在许多其他组织中(Hökfelt et al., 2003)。
与神经递质受体(微摩尔Kds)相比,神经肽受体具有相对较高的配体亲和力(纳摩尔Kds)。这样,少量扩散肽仍然可以激活受体。这一事实和它们的长寿命使神经肽在相对较低的浓度下在相对较长的距离上保持活性(Russo, 2017)。
差异表达与加工多样性
由于选择性剪接、串联组织或前肽的细胞特异性差异化翻译后加工,单个神经肽基因通常表现出多种表型(Albrechtsen和Rehfeld,2021)。当发现降钙素基因产生编码降钙素肽或降钙素基因相关肽(CGRPs)的mRNA时,选择性剪接被发现(Amara et al.1982)。对于某些神经肽,前肽被差异化加工以产生不同长度的成熟肽,这些肽释放出与受体结合的相同表位(图3A)(Rehfeld et al.2008)。虽然相同前体的不同产物与相同的受体结合,但它们从循环中清除的差异影响了它们的作用。因此,proCCK是否主要加工为CCK-33、CCK-12或CCK-8,或者prosomatostatin是否加工为生长抑素-28或生长抑素-14,这都是相关的(Albrechtsen和Rehfeld, 2021)。
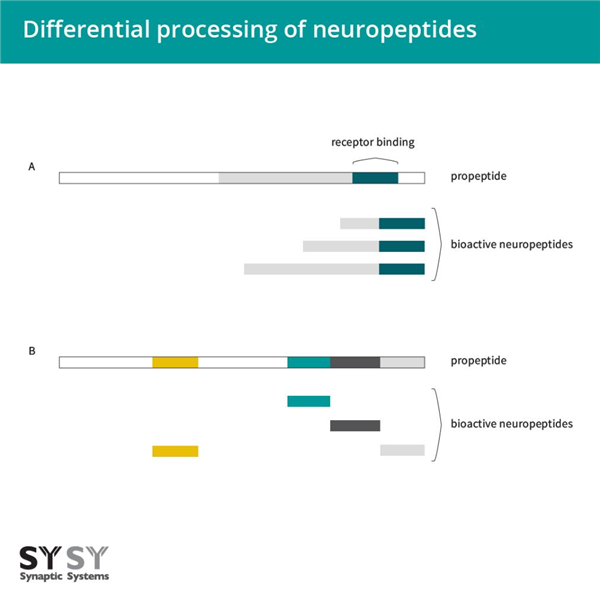
图3:神经肽的差异加工。 A:对于一些神经肽,前体肽的差异加工导致长度不同的成熟肽,但它们仍然共享相同的受体结合表位。 B:在某些情况下,前体肽包含不同的神经肽,可以在不同的组织中进行差异加工。
基因表达不同生物活性肽的另一种方式是基因本身编码含有不同神经肽的前肽(图3B),例如阿片肽基因和一些速激肽基因。 这些不同的神经肽存在于同一个前肽中,可以在不同的组织中进行差异化处理(Albrechtsen和Rehfeld,2021)。
多肽的功能
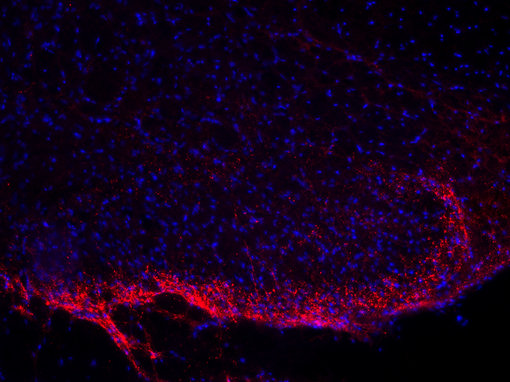
神经肽在多种靶组织中起作用,其作用可以是局部的或远距离的,因此几乎所有的身体功能都可以被调节(Russo, 2017)。许多具有相似结构的神经肽具有非常不同的功能。例如,加压素和催产素是两种下丘脑肽,每种都由9个氨基酸组成(见图4A和4B的脑切片的免疫组化染色)。这两种肽在其中7个残基上是相同的,被认为是进化早期基因重复的结果。这两种肽的作用是不同的:催产素引起乳汁分泌和子宫收缩,而加压素引起肾脏水潴留和血管收缩(Mains和Eipper,1999)。
 |
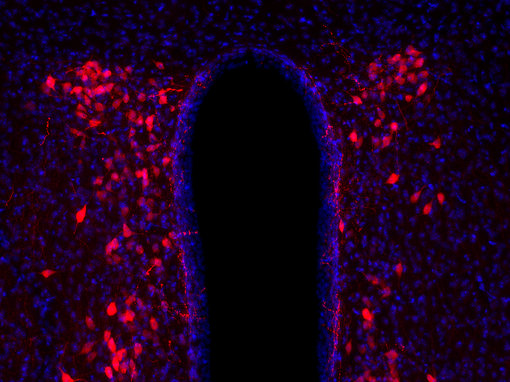 |
| 图4A:用豚鼠抗血管加压素抗体(cat. no. 403 004, dilution 1:500,红色)对PFA固定的大鼠下丘脑切片进行间接免疫染色。 通过DAPI染色(蓝色)观察到细胞核。 | 图4B:用豚鼠抗催产素抗体(cat. no. 408 004, dilution 1:500,红色)对PFA固定小鼠下丘脑切片进行间接免疫染色。 通过DAPI染色(蓝色)可观察到细胞核。 |
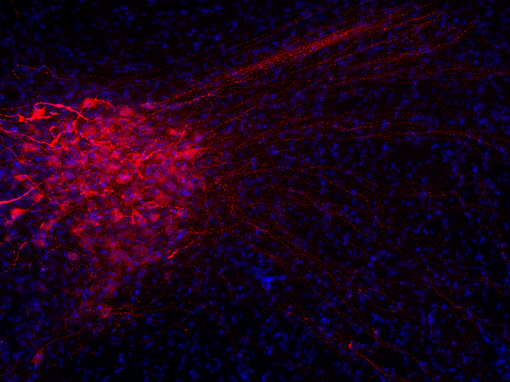
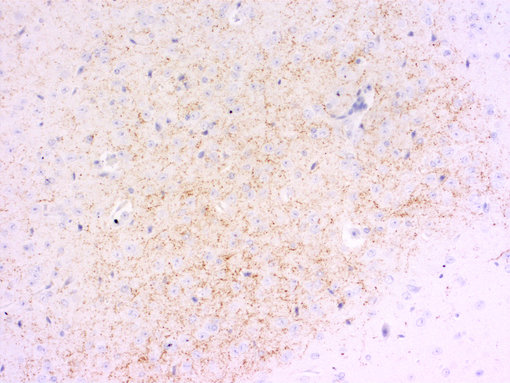
神经肽一直以来都对疼痛传递感兴趣。对转基因小鼠的研究表明,缺乏P物质或其受体的小鼠对中度或重度疼痛没有反应(Hökfelt et al., 2003)。另一种肽,降钙素基因相关肽(CGRP),在偏头痛的病理生理学中发挥重要作用(参见图5A和5B的脊髓切片染色)(Edvinsson et al., 2018, Russo, 2015)。
 |
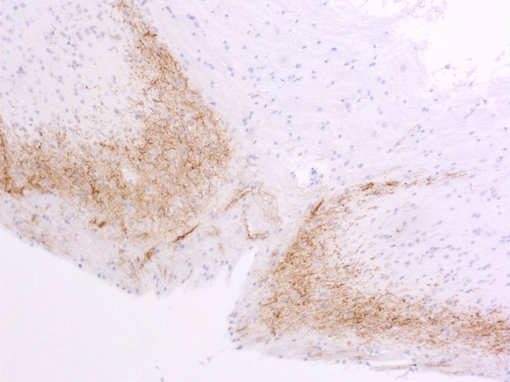 |
| 图5A:用豚鼠抗Substance P抗体(cat. no. 459 005,稀释度1:500,红色)对PFA固定大鼠脊髓切片进行间接免疫染色。 通过DAPI染色(蓝色)可观察到细胞核。 | 图5B:用豚鼠抗CGRP抗体(目录编号414 004,稀释度1:1000,DAB)对PFA固定石蜡包埋大鼠脊髓切片进行间接免疫染色。细胞核用苏木精(蓝色)复染。 |
中枢神经系统对食物摄入的控制是另一个正在进行的研究课题。神经肽Y刺激碳水化合物摄入,而半乳糖胺刺激脂肪摄入(见图6A和6B脑切片的免疫组化染色)。刺参相关肽和食欲素也有刺激作用。其他神经肽,如黑皮质素和可卡因-安非他命调节转录本,抑制食物摄入(Hökfelt et al., 2003)。
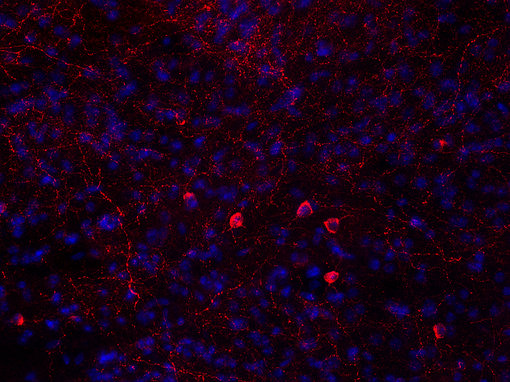 |
 |
| 图6A:用鸡抗神经肽Y抗体(cat. no. 394 006,稀释度1:500,红色)对PFA固定小鼠纹状体切片进行间接免疫染色。DAPI染色(蓝色)显示细胞核。染色前应用抗原提取(10mM Tris,1mM EDTA,pH 9.0,60°C过夜)。 | 图6B:用豚鼠抗Galanin抗体(目录编号446 004,稀释度1 : 1000,DAB)对PFA固定石蜡包埋小鼠下丘脑切片进行间接免疫染色。细胞核用苏木精(蓝色)复染。 |
总之,神经肽的各种功能与这组信号分子本身一样多样。以下是一些神经肽及其功能的列表:
| 神经肽 | 功能 |
| ACTH | 促进皮质醇的产生和释放 (Gallo-Payet, 2016) |
| AGRP | 刺激食欲,调节新陈代谢和能量消耗 (Ilnytska and Argyropoulos, 2008) |
| CART | 调节进食、奖赏和应激,作为精神兴奋剂 (Rogge et al., 2008) |
| CCK-8 | 参与消化、食物摄入、焦虑和恐惧 (Lee and Soltesz, 2011) |
| CGRP | 作为血管扩张剂和传递痛觉 (Benarroch, 2011) |
| CRF | 刺激ACTH的产生,决定妊娠长度和分娩时间 (Vitoratos et al., 2006) |
| Galanin | 参与调节进食、渗透压稳态、痛觉传导、觉醒/睡眠和认知 (Lang et al., 2015) |
| Neuropeptide S | 参与调节觉醒、焦虑和恐惧、食物摄入、学习和记忆 (Grund and Neumann, 2019) |
| Neuropeptide Y | 参与食物摄入、能量储存、减轻压力、焦虑和疼痛感知、调节血压 (Reichmann and Holzer, 2016) |
| Neurotensin | 调节多巴胺通路、疼痛、体温、食欲、脂肪代谢和学习和记忆 (Saiyasit et al., 2018) |
| Orexin | 调节进食、睡眠、觉醒和能量稳态 (Nixon et al., 2015) |
| Oxytocin | 刺激产后平滑肌收缩和泌乳,参与社会联系和繁殖 (Lee et al., 2009) |
| Somatostatin | 内分泌激素分泌的负调节因子 (Gehete et al., 2010) |
| Substance P | 肠道平滑肌收缩、血管扩张、中枢疼痛处理、神经源性炎症、焦虑和压力 (Schank and Heilig, 2017) |
| Vasopressin | 调节水分平衡、血压和社会行为 (Caldwell et al., 2008) |
| VIP | 刺激心脏收缩、血管扩张、调节血压和放松气管、胃和胆囊的平滑肌 (Iwasaki et al., 2019) |
疾病与药物开发
大量的神经肽和神经肽受体为药物靶点的发现提供了许多机会。食欲素神经元仅位于侧下丘脑,并支配大脑的广泛区域(见图7A和7B的免疫组化图片)。 几家制药公司正以这些系统为靶点,开发治疗肥胖症的药物(Hökfelt et al., 2003)。
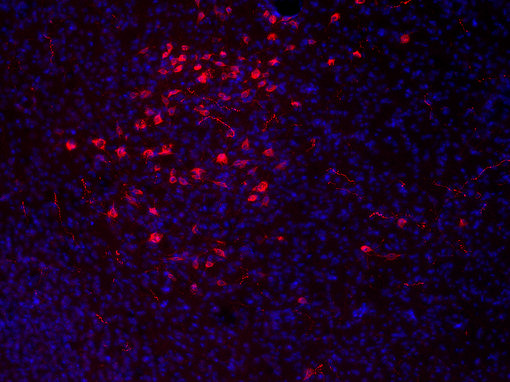 |
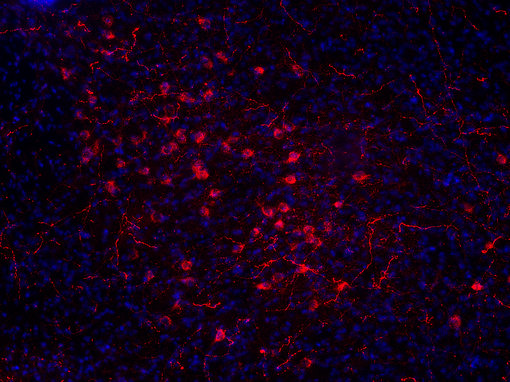 |
| 图7A:用豚鼠抗食欲素A抗体(目录编号389 004,稀释度1:500,红色)对PFA固定小鼠下丘脑切片进行间接免疫染色。 通过DAPI染色(蓝色)可观察到细胞核。 | 图7B:用豚鼠抗食欲素A/B抗体(目录编号389 104,稀释度1:500,红色)对PFA固定小鼠下丘脑切片进行间接免疫染色。 通过DAPI染色(蓝色)可观察到细胞核。 |
在P物质发现70年后,第一种肽类药物,一种P物质拮抗剂,被用于临床试验治疗抑郁症。肽类研究进展缓慢部分原因是合成选择性和强效的血脑屏障穿透激动剂或拮抗剂的困难(Hökfelt et al., 2003)。近年来,针对CGRP及其受体的单克隆抗体作为一种新的偏头痛治疗方法被引入,它们通过阻断CGRP信号通路来预防偏头痛,是目前最先进的技术(Sevivas和Fresco,2022年,Vandervorst等,2021年)。
靶向神经肽的抗体
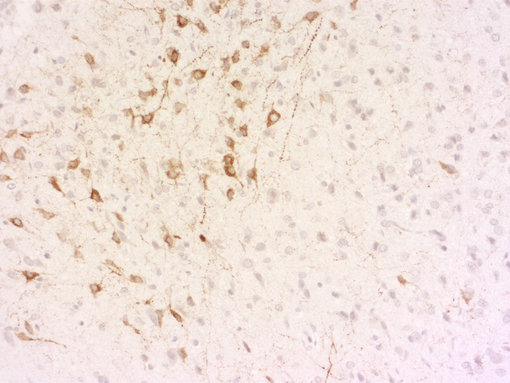
一般而言,我们的目标是开发针对活性肽的抗体,并使用加工过的肽或切割活性肽的末端部分,包括已知的修饰,用于免疫。我们的抗体在免疫组化(IHC和IHC-P,如图8A)或免疫细胞化学(ICC,图8B)中表现出优异的性能,可以作为您研究的有价值的实验试剂!
 |
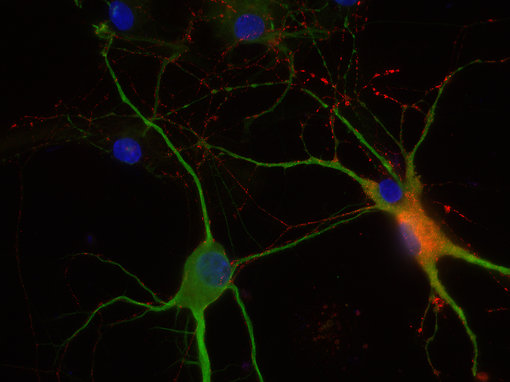 |
| 图8A:用豚鼠抗生长抑素-28抗体(cat. no. 366 004, dilution 1:500, DAB)对PFA固定石蜡包埋的大鼠下丘脑切片进行间接免疫染色。 通过苏木精染色(蓝色)可观察到细胞核。 | 图8D:用豚鼠抗VIP抗体(目录编号443 005,稀释度1:100,红色)和兔抗MAP 2抗体(目录编号188 002,稀释度1:1000,绿色)对PFA固定的大鼠海马神经元进行间接免疫染色。 通过DAPI染色(蓝色)可观察到细胞核。 |
神经肽和肽类激素
| 货号 | 产品名称 | 应用类型 | 规格 |
| 452 005 | ACTH, Guinea pig, polyclonal, affinity purified | IHC IHC-P | 50ug |
| 438 004 | CCK-8, Guinea pig, polyclonal, antiserum | ICC IHC IHC-P | 100ul |
| 414 004 | CGRP, Guinea pig, polyclonal, antiserum | IHC IHC-P iDISCO Clarity | 100ul |
| 259 003 | Chromogranin A, rabbit, polyclonal, affinity purified K.O. | WB ICC IHC IHC-P | 50ug |
| 259 002 | Chromogranin A, rabbit, polyclonal, antiserum | WB | 200ul |
| 259-0P | Chromogranin A, control protein | 100ug | |
| 259 103 | Chromogranin B, rabbit, polyclonal, affinity purified K.O. | WB ICC IHC IHC-P | 50ug |
| 259-1P | Chromogranin B, control protein | 100ug | |
| 446 004 | Galanin, Guinea pig, polyclonal, antiserum | ICC IHC IHC-P | 100ul |
| 468 003 | Ghrelin, rabbit, polyclonal, affinity purified | IHC IHC-P | 50ug |
| 460 003 | GIP, rabbit, polyclonal, affinity purified | IHC IHC-P | 50ug |
| 471 005 | GLP-1, Guinea pig, polyclonal, affinity purified | Dot blot IHC IHC-P | 50ug |
| 434 005 | Neuropeptide S, Guinea pig, polyclonal, affinity purified | IHC IHC-P | 50ug |
| 394 006 | Neuropeptide Y, chicken, polyclonal, affinity purified K.O. | ICC IHC | 200ul |
| 394 004 | Neuropeptide Y, Guinea pig, polyclonal, antiserum | ICC IHC IHC-P | 100ul |
| 418 005 | Neurotensin, Guinea pig, polyclonal, affinity purified | IHC IHC-P | 50ug |
| 389 004 | Orexin A, Guinea pig, polyclonal, antiserum K.O. | IHC IHC-P | 100ul |
| 389 104 | Orexin A/B, Guinea pig, polyclonal, antiserum | IHC IHC-P | 100ul |
| 408 004 | Oxytocin, Guinea pig, polyclonal, antiserum K.O. | IHC IHC-P | 100ul |
| 366 006 | Somatostatin-28, chicken, polyclonal, affinity purified | ICC IHC IHC-P | 200ul |
| 366 004 | Somatostatin-28, Guinea pig, polyclonal, antiserum | ICC IHC IHC-P | 100ul |
| 366 017 | Somatostatin-28, rat, monoclonal, purified IgG | ICC IHC IHC-P | 100ug |
| 459 005 | Substance P, Guinea pig, polyclonal, affinity purified | IHC | 50ug |
| 403 004 | Vasopressin, Guinea pig, polyclonal, antiserum | IHC IHC-P | 100ul |
| 443 005 | VIP, Guinea pig, polyclonal, affinity purified | ICC IHC IHC-P | 50ug |
原作者:SySy--Dr. Beate Friedrich
 |
贝亚特拥有深厚的生物化学背景,负责重组抗体部门。她对神经肽和肽激素特别感兴趣,并负责该产品组的抗体开发。 |
参考文献
- Albrechtsen and Rehfeld, 2021: On premises and principles for measurement of gastrointestinal peptide hormones.PMID: 33811948IF: 3.0 Q3
- Amara et al., 1982: Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. PMID: 6283379IF: 64.8 Q1
- Benarroch, 2011: CGRP: sensory neuropeptide with multiple neurologic implications. PMID: 21768598IF: 9.9 Q1
- Caldwell et al., 2008: Vasopressin: behavioral roles of an “original” neuropeptide. PMID: 18053631IF: 6.7 Q1
- Edvinsson et al., 2018: CGRP as the target of new migraine therapies – successful translation from bench to clinic. PMID: 29691490IF: 38.1 Q1
- Hökfelt et al., 2003: Neuropeptides: opportunities for drug discovery. PMID: 12878434IF: 48.0 Q1
- Hook, 2008: Proteases for processing proneuropeptides into peptide neurotransmitters and hormones. PMID: 18184105IF: 12.5 Q1
- Ilnytska and Argyropoulos, 2008: The role of the Agouti-related protein in energy balance regulation. PMID: 18470724IF: 8.0 Q1
- Kastin, 2000: What is a neuropeptide? PMID: 10675912IF: 15.9 Q1
- Lang et al., 2015: Physiology, signaling, and pharmacology of galanin peptides and receptors: three decades of emerging diversity. PMID: 25428932IF: 21.1 Q1
- Nixon et al., 2015: Sleep disorders, obesity, and aging: the role of orexin. PMID: 25462194IF: 13.1 Q1
- Rogge et al., 2008: CART peptides : regulators of body weight, reward and other functions. PMID: 18802445IF: 34.7 Q1
- Russo, 2015: Calcitonin gene-related peptide (CGRP): A new target for migraine. PMID: 25340934IF: 12.5 Q1
- Van den Pol, 2012: Neuropeptide transmission in brain circuits. PMID: 23040809IF: 16.2 Q1
更多神经肽与SySy抗体产品,欢迎垂询SySy中国区金牌代理与畅货中心,艾美捷科技,400-6800-868,www.amyjet.com